

/image%2F0550795%2F201304%2Fob_ae1487_2012-06-03-fukushima-photo-jm-bonzom-010.jpg)
Chronic sublethal stress causes bee colony failure
8 Novembre 2013 , Rédigé par Bioécologie Publié dans #Faune et Flore, #Biodiversité, #Chercheurs - projets de recherche..., #Livres - revues - thèses - rapports..., #Actualités-News
John Bryden, Richard J. Gill, Robert A. A. Mitton, Nigel E. Raine and Vincent A. A. Jansen, Ecology Letters, (2013) 16:1463–1469
Abstract
Current bee population declines and colony failures are well documented yet poorly understood and no single factor has been identified as a leading cause. The evidence is equivocal and puzzling: for instance, many pathogens and parasites can be found in both failing and surviving colonies and field pesticide exposure is typically sublethal. Here, we investigate how these results can be due to sublethal stress impairing colony function. We mathematically modelled stress on individual bees which impairs colony function and found how positive density dependence can cause multiple dynamic outcomes: some colonies fail while others thrive. We then exposed bumblebee colonies to sublethal levels of a neonicotinoid pesticide. The dynamics of colony failure, which we observed, were most accurately described by our model. We argue that our model can explain the enigmatic aspects of bee colony failures, highlighting an important role for sublethalstress in colony declines.
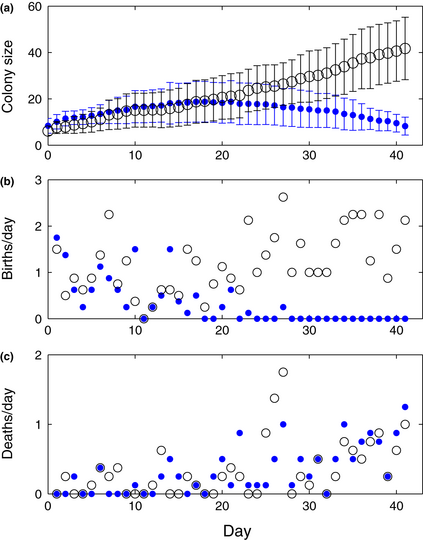
Figure 2. Comparison of neonicotinoid treated colonies (blue filled circles) with control colonies (black open circles). (a) Both treatment and control (plot shows mean numbers of workers ± 95% CI) have similar growth at first, but trajectories begin to diverge after approximately 3 weeks. (b, c) Worker birth and death rates (mean number of births and deaths per colony are shown) for treatment and control colonies illustrate the impact of the pesticide on colony health. In the treatment colonies, the birth rate decreases while colony size increases during the first 21 days, and after 21 days the death rate increases while colony size decreases.
See the attached file tfor the publication
Ici, une présentation en français de ce travail, par Stéphane Foucart, journaliste au journal Le Monde
Article first published online: 6 OCT 2013



/image%2F0550795%2F20181020%2Fob_2ce219_1-poisson-yoanna-bd-laureate.jpg)
/image%2F0550795%2F20180321%2Fob_75f43e_bios-1650270-72dpi.jpg)
/image%2F0550795%2F20180215%2Fob_aaac89_944453-113585545510761-27780121-n.jpg)
/idata%2F2352969%2Fphotos%2Fpeacock-spider-maratus-species-a-peckhamania-photo-Jurgen-.jpg)
Suivez-moi
Newsletter
Abonnez-vous pour être averti des nouveaux articles publiés.
Catégories
- 205 A voir - à écouter
- 181 Actualités-News
- 178 Livres - revues - thèses - rapports...
- 174 Photos - arts - webdocumentaires...
- 154 Faune et Flore
- 154 Thèses - Post-doc - Emploi - Bourse
- 132 Biodiversité
- 100 Statistique - modélisation - plans expérimentaux
- 89 Matériels et Méthodes
- 83 Chercheurs - projets de recherche...
- 77 Ecotoxicologie
- 43 Océanographie
- 35 Evaluation des risques et surveillance écologiques
- 33 Radionucléides - chimiques - nanoparticules...
- 32 Colloques - conférences
- 28 Organismes de laboratoire
- 23 Spécial Japon
- 22 Cours...
- 21 Evolution
- 21 Universités - Laboratoires... : focus !
- 18 A voir
- 15 Bioturbation
- 15 Etudiant(e)s : conseils
- 13 Amphibiens
- 12 Logiciels scientifiques - utilitaires gratuits
- 11 Sociétés savantes - alliances...
- 8 Sédiment: biogéochimie et bioturbation
- 7 projets de recherche...
- 5 Photos
- 3 Emploi
- 3 Etudiant(e)s : conseils
- 3 In Memoriam
- 2 Bourse
- 2 Chercheurs
- 2 Post-doc
- 2 Sédiment: biogéochimie et bioturbation
- 2 Thèses
- 2 Universités - Laboratoires... : focus !
- 2 arts
- 2 webdocumentaires
- 2 à écouter
- 1 ...
- 1 Statistique
- 1 e
- 1 sol
